Molecular Basis of Hearing
Hearing: Turning Sound into Voltage
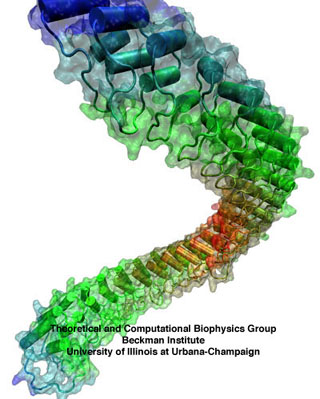 Ankyrin, a soft spring in the inner ear.
Ankyrin, a soft spring in the inner ear.
The ear is a sensitive and robust device, able to perceive the faint sound of flowing water and the thunderous blast of an air plane. Like a microphone, the ear transforms a complex, mechanical stimulus (sound), into an electrical signal, a voltage change in a nerve cell, that can be understood by our brain. This transformation is called "mechanotransduction" and is accomplished by a series of amazingly minute devices that each connect a soft spring to an ion channel, both located in specialized sensory cells, the hair cells of the inner ear. The springs, through their vibrations agitated by particular sound frequencies, control ion currents passing through the channels, thereby, modifying the hair cell internal electrical potential. This leads to neural signaling to the acoustic cortex of the brain. Recently reported molecular dynamics simulations using NAMD, some of the most extensive simulations accomplished to date both in size and duration, showed that the mechanical characteristics of hair cell signaling might be traced to a single protein, ankyrin, that acts as a helical spring. Imagine a soft spring that is extended several inches by the weight of a feather! Ankyrin is such a spring, but a billion times finer, and found in the inner ear. It's role in hearing is not established yet, but even if it is not involved in hair cell mechanotransduction, its remarkable elastic properties (first predicted using molecular dynamics and later confirmed through AFM experiments) are likely relevant for other cellular processes.
Hair Cells and Molecules of Mechanotransduction
The vertebrate senses of hearing and balance employ mechanically sensitive hair cells to transform complex mechanical stimuli produced by sound and acceleration into electrical signals. In both the auditory and vestibular systems, these stimuli bend the hair cells' stereocilia, microvillus-like organelles that project in bundles from the apical surfaces of the cells, and the bending opens mechanically gated transduction channels in the tips of stereocilia to allow ion influx. The biophysics of this process is well understood, and sensitive mechanical measurements have characterized the force needed to open channels and the stiffness of structures that convey this forces. The microscopic structures involved are also well understood: Stereocilia in each hair bundle are arranged in rows of increasing height, and a fine filament termed a tip link connects the tip of each stereocilium to the side of its taller neighbor. Tip links are thought to connect directly to transduction channels, so that deflections of a hair bundle that would tighten tip links open these channels. Proteins forming the tip link are unknown, but cadherin 23, a long cadherin with 27 extracellular domains and related to hereditary deafness, is present in stereocilia and may be part of the tip links or transient lateral links of hair cells. The transduction channel is likely to contain subunits of the TRP channel family, perhaps TRPN1 (NOMPC) in lower vertebrates and TRPA1 (ANKTM1) in all vertebrates. Both have extended N-termini with 29 or 17 ankyrin repeats, respectively. In order to have an idea of the size of these molecules, imagine that a finger (~1 cm wide) is a stereocilium. Then, a human hair would be ~1 m wide and a stack of ankyrin repeats would be 0.04 mm thick!
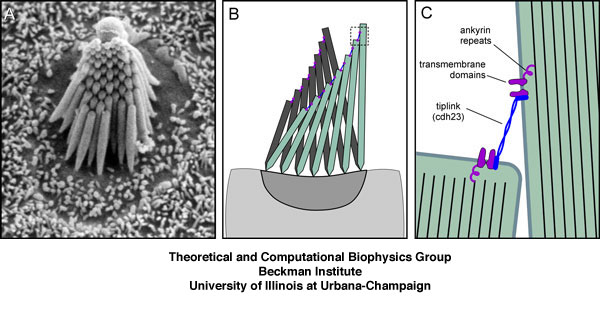
Mechanotransduction in hair cells of the inner ear. (A) Scanning electron micrograph of hair bundle (bullfrog sacculus; David P. Corey's Lab.). This top view shows the stereocilia arranged in order of increasing height. (B) Model for mechanotransduction. Deflection of a hair cell's bundle causes the stereocilia to bend and the tip links between them to tighten. (C) Ion channels attached to intracellular elastic elements (ankyrin repeats) open in response to tension on the rather inextensible tip link.
Cadherin: The Stiff String
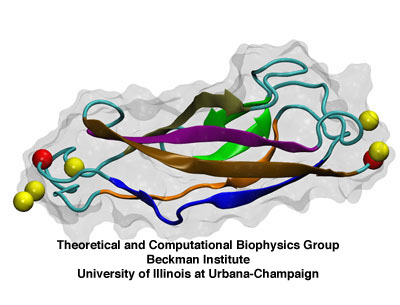 Single extracellular cadherin repeat. The seven beta-strands are shown
as thick arrows and colored blue, orange, tan, green, purple, and
ochre. C_alpha atoms of the termini are shown as red spheres and calcium
ions as yellow spheres.
Single extracellular cadherin repeat. The seven beta-strands are shown
as thick arrows and colored blue, orange, tan, green, purple, and
ochre. C_alpha atoms of the termini are shown as red spheres and calcium
ions as yellow spheres.
Cadherin proteins which play a fundamental role in calcium-dependent cell-cell adhesion are typically formed by a single-pass transmembrane domain and multiple extracellular repeats. In general, each cadherin repeat consists of about 110 amino-acids sharing a common folding topology characterized by seven antiparallel beta-strands tightly linked by hydrogen bonds and a highly conserved calcium binding motif. The microscopic architecture of cadherins along with high resolution electron micrographs suggest that the tip link is actually a rather stiff element. Steered molecular dynamics (SMD) have shown that indeed individual cadherin domains are very stiff for small forces, and that forces of hundreds of picoNewtons cause their complete unfolding. In fact, the magnitude of the force peaks observed when stretching single cadherin domains, with or without calcium ions, is significantly larger (by a factor of >10) than the force required to stretch or unfold multiple ankyrin repeats under the same simulation conditions.
- Click here for a movie (mpeg, 4.0M) showing unfolding of a single cadherin domain. The role of calcium ions as mechanical stabilizers becomes evident in the simulation.
Ankyrin: The Soft Spring
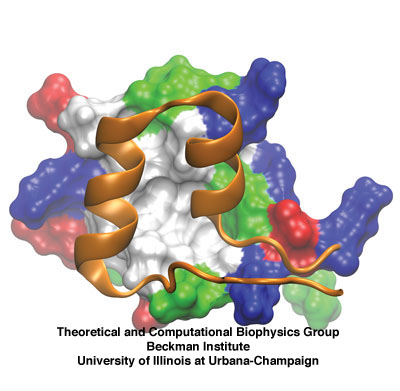 Side view of two ankyrin repeats. The two alpha-helices of one ankyrin
repeat are shown in cartoon representation (orange). Behind, another
repeat is shown in surface representation and colored by residue type.
Side view of two ankyrin repeats. The two alpha-helices of one ankyrin
repeat are shown in cartoon representation (orange). Behind, another
repeat is shown in surface representation and colored by residue type.
Ankyrin repeats are 33-amino-acid domains that occur in sets of four or more, and appear in more than 400 human proteins expressed in many tissues. Each repeat has two short antiparallel alpha-helices and a less ordered loop, with the helix pairs stacked in parallel in adjacent repeats. Proteins of the ankyrin family, for which the repeat was named, contain up to 24 repeats. The largest ankyrin-repeat crystallographic structure available to date corresponds to 12 of the repeats from the human ankyrin-R, and a structure with 24 repeats has been extrapolated from it. There is a slight curvature to the stacking, so that 25-30 repeats form a super-helical structure with a complete turn as shown below. Different ankyrin repeats within a single protein or in different proteins have a rather conserved, but not identical amino-acid sequence. A less-conserved loop linking adjacent repeats mediates protein-protein interactions in some proteins with ankyrin repeats.
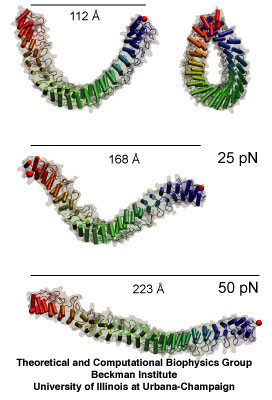 Elastic response of 24 ankyrin repeats. Snapshots of the equilibrated
structure (front and side views); the structure stretched by a force of 25
pN over 10 ns; and the structure stretched by a force of 50 pN over 10 ns.
Elastic response of 24 ankyrin repeats. Snapshots of the equilibrated
structure (front and side views); the structure stretched by a force of 25
pN over 10 ns; and the structure stretched by a force of 50 pN over 10 ns.
Steered molecular dynamics simulations have shown that stacks of 12-24 ankyrin repeats form a molecular spring that is reversible on a nanosecond time scale and that has a stiffness very close to that of the "gating spring" (see "Molecualr basis of mechanosensory transduction", P.G. Gillespie and R. G. Walker, Nature 413:194-202, and references therin), an elastic element that pulls on transduction channels. A similar elastic response was observed for a model of 17 ankyrin repeats of TRPA1. In addition, forces just above the working range cause unpacking of helices to extend the protein by nearly ten-fold, which may serve as a safety release. The simulations suggest that the ankyrin repeats of TRPA1 and TRPN1 are involved in mechanical gating and may serve as the gating spring for hair-cells. While the latter hypothesis has been challenged by more recent experimental results, the unique elastic properties of ankyrin have been confirmed through AFM experiments as described below.
- Click here for a movie (mpeg, 2.8M) showing a model of 24 ankyrin repeats of human ankyrin-R followed by 1.2 ns of a constant force stretch simulation (100 pN).
- Click here for a movie (mpeg, 1.7M) based on a 25 ns relaxation of 24 ankyrin repeats of human ankyrin-R played back and forth.
- Click here for a movie (mpeg, 1.8M) showing a model of 17 ankyrin repeats of mmTRPA1 followed by 6.5 ns of a constant force stretch simulation (50 pN).
- Click here for a movie (mpeg, 2.8M) showing unfolding of 4 identical ankyrin repeats. Detachment and unfolding of individual helices of the third and last repeats can be recognized.
Theoretical Predictions Confirmed through AFM Experiments!
The mechanical response of ankyrin proteins is characterized at low-force by an elastic behavior reflecting changes of the overall shape (tertiary structure elasticity, TSE). Steered molecular dynamics simulations have also shown that at high-force the mechanical response of these proteins is characterized by elasticity stemming from sequential unraveling of secondary structure elements (secondary structure elasticity, SSE). Both tertiary and secondary structure elasticity of ankyrin repeats, predicted first through simulations, have been recently confirmed in AFM experiments. Indeed, a first report by Piotr Marszalek and collaborators shows that a stack of 24 ankyrin repeats from human ankyrinB behaves as a reversible, linear, nanospring, its properties based on TSE. The same report and an additional paper by Julio Fernandez and collaborators show that mechanical unfolding of ankyrin repeats occurs through a sequence of steps (SSE). These experimental results have been interpreted in light of prior steered molecular dynamics simulations calculations, which provide a unique microscopic view of ankyrin's elasticity.
Is there a Numerical Error Effect on Ankyrin Elastic Properties?
Classical molecular dynamics solves Newton's equations of motion for a system composed of many particles/atoms that interact through a given potential energy. The equations of motion are integrated using time-reversible algorithms that employ discrete time-steps. This leads to numerical errors, e.g., to an energy increase and, hence a temperature increase, as reported and briefly discussed in our published report (see citation below). This increase in temperature raised concerns about its effect on the simulated elastic response of ankyrin (Joe Howard, private communication). We have carried out additional simulations that decreased the integration time steps to rather impracticable (very long computing times) values. These control simulations yielded the same ankyrin elastic properties as reported above, while the increase in temperature was drastically reduced (less than one degree Celsius during six nanoseconds) and energy was better conserved (to better than 0.5% over six nanoseconds). The results of the new simulations are presented and discussed in detail here.
Publications
Tightrope act. David P. Corey and Marcos Sotomayor. Nature, 428, 901-902, 2004.
Investigators
Related TCB Group Projects
- Mechanosensitive Channel MscS
- Mechanosensitive Channel MscL
- Cells Get Sticky with Calcium
- Mechanical Proteins
Page created and maintained by Marcos Sotomayor.


