Structure, Dynamics, and Function of Aquaporins
 2004 Winner of
Visualization Challenge in Science and Engineering,
2004 Winner of
Visualization Challenge in Science and Engineering, Organized by the National Science Foundation and Science Magazine. (See the related Science article.)
Our water permeation movie and the 2003 Nobel prize for Chemistry
The Art of Water Transport in Aquaporins
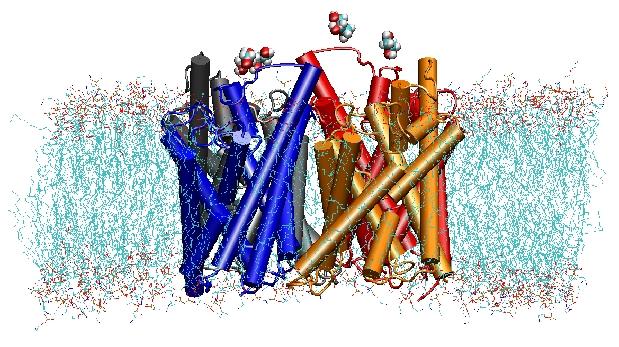
Aquaporins are membrane water channels that play critical roles in controlling the water contents of cells. These channels are widely distributed in all kingdoms of life, including bacteria, plants, and mammals. More than ten different aquaporins have been found in human body, and several diseases, such as congenital cataracts and nephrogenic diabetes insipidus, are connected to the impaired function of these channels. They form tetramers in the cell membrane, and facilitate the transport of water and, in some cases, other small solutes across the membrane. However, the water pores are completely impermeable to charged species, such as protons, a remarkable property that is critical for the conservation of membrane's electrochemical potential, but paradoxical at the same time, since protons can usually be transfered readily through water molecules. The results of our simulations have now provided new insight into the mechanism underlying this fascinating property. Water molecules passing the channel are forced, by the protein's electrostatic forces, to flip at the center of the channel (see the animation), thereby breaking the alternative donor-acceptor arrangement that is necessary for proton translocation (read the complete story in our Science paper).

Functional Implication of a Peculiar Protein Architecture
In the year 2000, Stroud's lab at UCSF succeeded in solving the first high resolution structure of an aquaporin by x-ray crystallography. The studied structure was the one of the E. coli glycerol uptake facilitator (GlpF), which is an aquaglyceroporin, i.e., the channel is also permeable to small linear sugar molecules such as glycerol. Nanosecond MD simulations of tetrameric GlpF in a hydrated patch of POPE lipid bilayer characterized the complete pathway of substrate conduction in the channel. Analysis of hydrogen bond interactions of the substrate with the interior of the channel also explained for the first time why these channels incorporate in their architecture two characteristic loops, including energetically unfavorable secondary structure elements, which are conserved in the whole aquaporin family (Jensen et al., Structure, 2001).
Energetics of Sugar Permeation
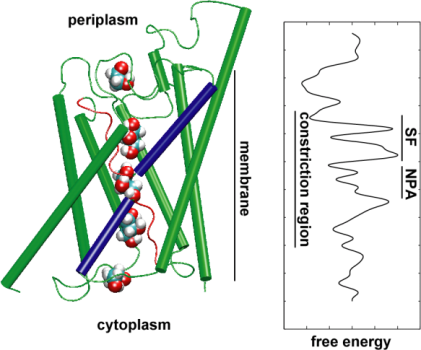
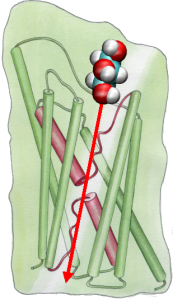
Then, we explored the energetics of the transport event to understand what barriers a substrate needs to overcome during its passage through the channel. In order to describe the free energy profile, we calculated the potential of mean force for the complete conduction event from the trajectories in which the movement of glycerol through the channel was accelerated by applying external forces. This technique, known as steered molecular dynamics (SMD), allows one to observe slow events within the accessible time scale of MD simulations. However, due to the non-equilibrium nature of the trajectories, in analyzing results one faces the problem of irreversible work that has to be discounted. Using Jarzinsky's identity, the equality between the free energy and work in non-equilibrium systems, we could completely describe the energetics of glycerol transport through the channel. The calculated potential of mean force captures major binding sites and barriers in the channel in close agreement with the results of equilibrium MD simulations and the crystal structure. Moreover, it displays a pronounced asymmetry in its shape, suggesting that the asymmetric structure of the protein may be functionally important for an efficient uptake of nutrient molecules from the environment. (Jensen, et al., PNAS, 2002).
What Makes an Aquaporin a Glycerol Channel
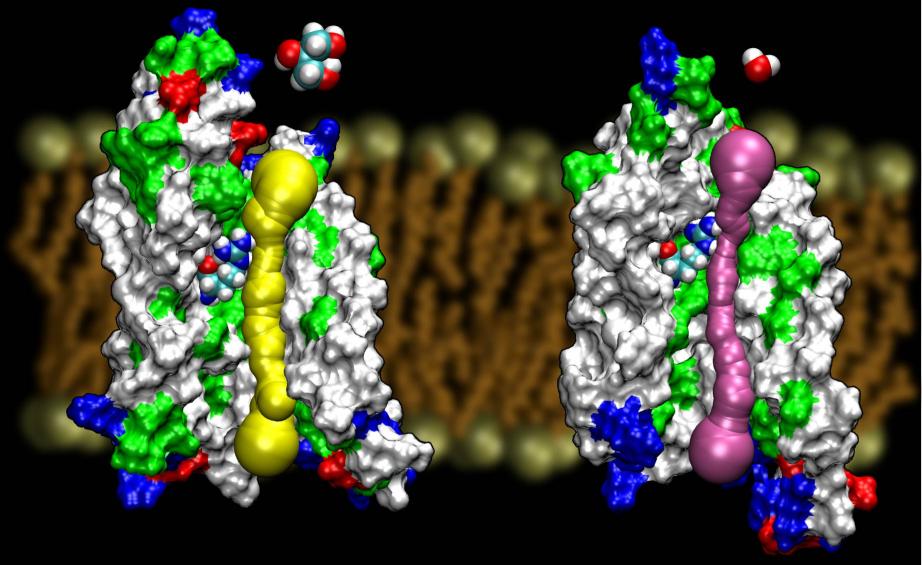
Aquaporins are a very lucky family of membrane proteins, which are notoriously resistant against structure determination. We have now high resolution structures of several AQPs: GlpF, AQP1, AQP0, and AqpZ. More structures are expected to emerge soon. The availability of the structure of two AQPs from E. coli, one that is a pure water channel (AqpZ) and the other one being a glycerol channel (GlpF) provided us with a unique opportunity to understand the selectivity principles employed by nature in using two genetically and structurally highly homologous channels for membrane trafficking of different materials. Comparison of the barriers calculated for conduction of glycerol in these two channels showed that AqpZ takes advantage of steric barriers to make it difficult for glycerol to go through. In fact, the pore in overall narrower than GlpF (see figure below). It seems that nature has turned to a very basic mechanism, namely tuning the pore size of the channel, in order to make two proteins from the same family exhibit different conduction properties. Read the entire story in Wang, et al., STRUCTURE, 2005.
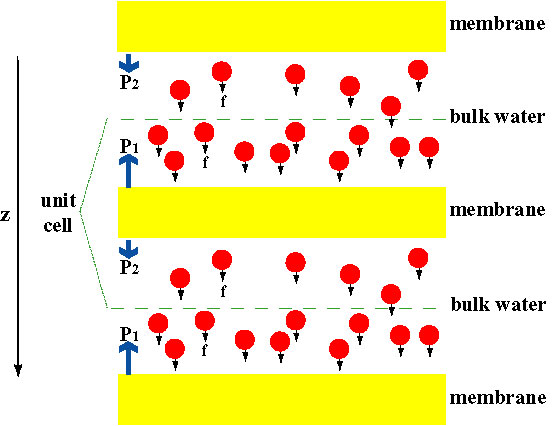
Putting Channels under Pressure
In order to directly compare the results of MD simulations with biochemical measurements of the conductivity of membrane water channels, in which osmotic pressure gradients are used to induce the flow of water across the membrane, we developed a new methodology for MD simulations. By applying small forces on water molecules in the bulk region (see figure), a hydrostatic pressure gradient is generated across the membrane. The pressure gradient induces conduction rates that can be studied with MD simulations, which are usually limited to a few nanoseconds. The method has been applied in the simulation of water permeation through the GlpF, and has resulted in a linear correlation between the applied pressure and the flux (Zhu et al., Biophys. J., 2002).
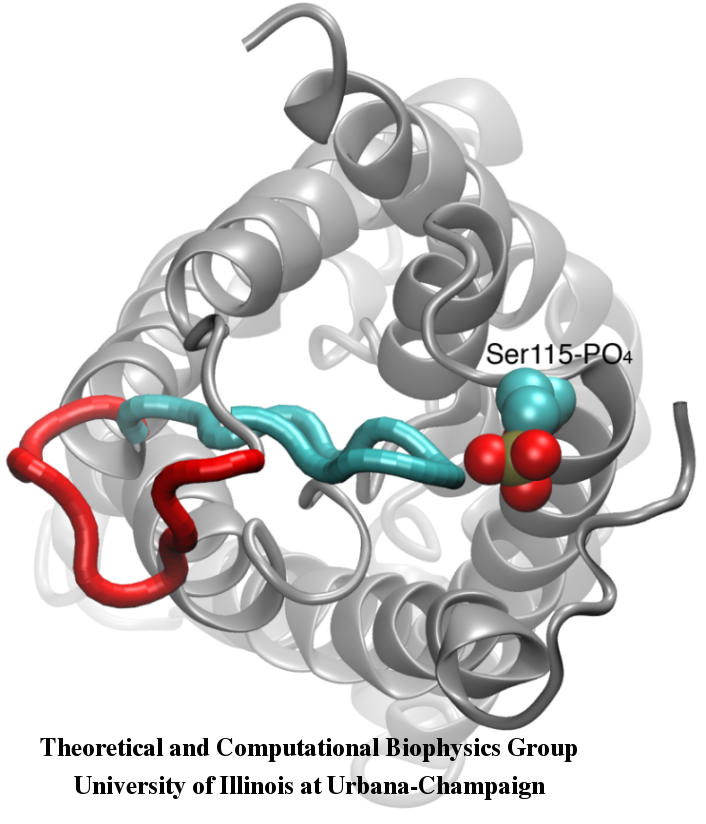
Gating of Water Channels
Water permeation through aquaporins is a passive process that follows the direction of osmotic pressure across the membrane. Although many aquaporins function as always-open channels, a subgroup of aquaporins, particularly in plants have evolved a sophisticated molecular mechanism through which the channel can be closed in response to harsh conditions of the environment, under which exchange of water can be harmful for the organism. Examples of such conditions are drought stress and flooding, which trigger certain cellular signals (dephosphorylation and change of pH) that result in closure of the channel. A recently solved structure of an aquaporin from spinach was simulated under different conditions to investigate the molecular mechansim of gating in these membrane channels. It was found that an elongated cytoplasmic loop in gating-competent aquaporins physically occludes the entrance of the water pore in the closed channel and blocks the access of water molecules to the pore. Displacement of this loop in response to phosphorylation, as observed in simulations, removes the loop from the cytoplasmic entrance of the pore. Furthermore, a hydrophobic amino acid which is conformationally closely coupled to this loop will be pulled back from the channel as a result of the movement of the loop. Together, these molecular events result in an open channel that is permeable by water molecules. The study appeared in Nature 2006.
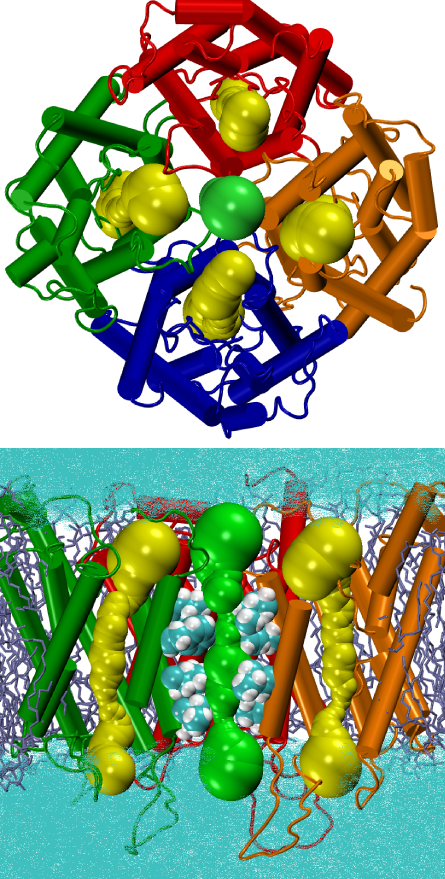
Gating and Ion Conductivity of the Tetrameric Pore
AQPs are generally known for their high permeability to water. However, their involvement in other cellular functions, including permeation of small molecules other than water, as well as cell-cell communication has been suggested. Tetramerization is a common structural feature of AQPs. The fact that the four monomers each form a functionally independent water pore, but require a tetrameric organization for function, suggests a synergistic benefit of oligomerization, and, thus, compels further analysis of the potential role of the central pore in AQPs. An interesting proposition is the possible involvement of the central pore in ion conduction upon cGMP activation. Using MD simulations of membrane embedded models of AQP1, we have investigated this property of AQP1, to determine how, and under what conformational and hydration conditions, the central pore might conduct ions and how the conductivity is regulated. One of the cytoplasmic loops, namely the arginine-rich loop D, was suggested by our simulations to play a key role in cGMP-mediated activation of the central pore. The suggested role of loop D was verifired experimentally by a designed mutant of loop D showing a complete loss of ion conductivity ( Yu et al., Structure, 2006 ) .
Gas Molecules Commute into Cell
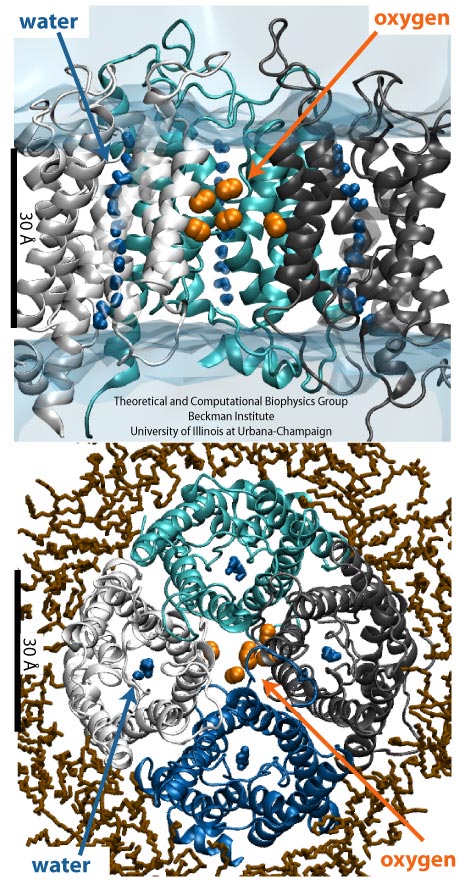
Every morning, many people drive to work, while others may bike, take the bus or the metro. Similarly, various biomolecules in the human body also reach their destinations in diverse manners. For example, to cross the cellular membrane, small hydrophobic gas molecules diffuse through the lipid bilayer, while water molecules pass through aquaporins (AQPs). Interestingly, just like one may get to work both by bus and by driving, it has been found recently that some gas molecules may have more than one way to cross the membrane, i.e., besides diffusion through lipids, oxygen and carbon dioxide may also pass through AQPs. However, the pathways that these gas molecules take remained elusive. Using molecular dynamics performed with NAMD, we have investigated the gas permeability of AQP1 with two complementary methods (explicit gas diffusion simulation and implicit ligand sampling). The simulation results suggest that while the four monomeric pores of AQP1 function as water channels, the central pore of AQP1 may serve as a pathway for gas molecules to cross the membrane (Wang et al., Journal of Structural Biology, 2007 ).
Molecular Obstacle Course
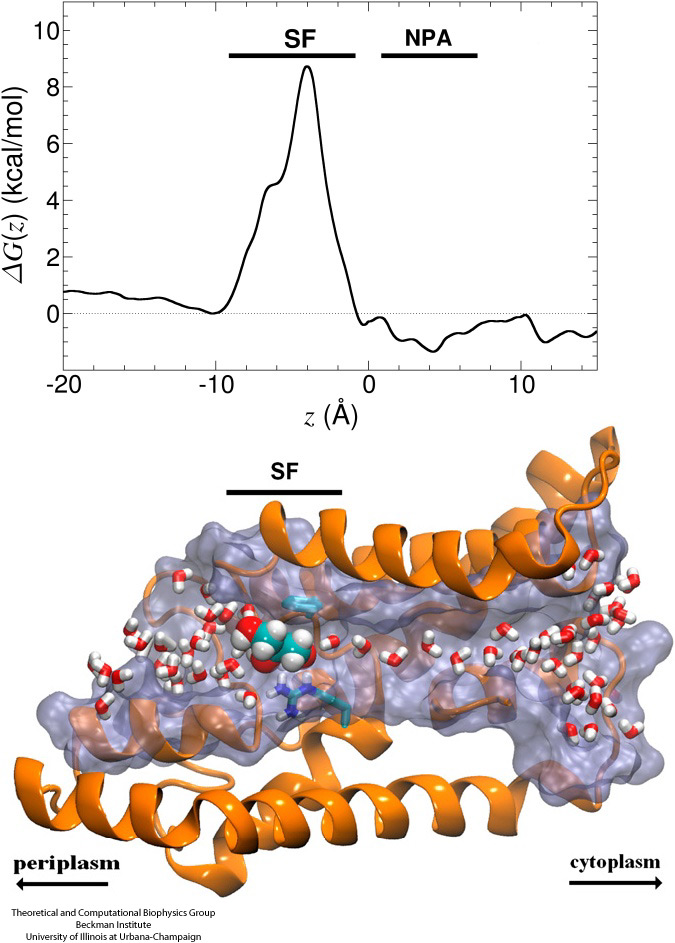
Biological cells protect their interior through their cellular membranes, yet rely on import of nutrients. They have evolved for this import fast conduction channels that include reliable checkpoints distinguishing desirable and undesirable compounds. A checkpoint puts up a veritable obstacle course that only the right compounds can pass quickly. Understanding the channel design is difficult due to lack of detailed experimental data on nutrient dynamics. Presently, the most detailed information comes from viewing channel dynamics computationally, starting from static crystallographic structures. A recent study investigated how glycerols, small nutrient molecules needed by some bacteria, pass through checkpoints realized through glycerol channel GlpF. GlpF furnishes four parallel channels that were monitored computationally using NAMD and a novel algorithm that explores the channel energetics quickly enough to be methodologicaly feasible on today's computers. The results show how the physical characteristics of glycerol, their ability to form so-called hydrogen bonds, their electrical dipole moments, their diffusive mobility and their intrinsic flexibility are probed along the channel (Henin et al., Biophysical Journal, in press ).
Publications
Molecular Basis of Proton Blockage in Aquaporins. Nilmadhab Chakrabarti, Emad Tajkhorshid, Benoît Roux, and Régis Pomès. Structure, 12: 65-74, 2004.
Last updated: Ocy 15, 2006 by Emad Tajkhorshid


