Lipid Signaling Controlled by PI4K Kinase Palmitoylation
Role of kinase in lipid signaling
Interplay between cellular membranes and their peripheral proteins drives many cellular processes, including cell division, growth, movement and cell-cell communication. Membrane lipids, which are long considered as passive structural molecules, can also act as signaling molecules to provide location-specific and temporal-controlled signaling. Lipid signaling is controlled by kinases and phosphatases via reversible phosphorylation as on-off switches. One type of kinases, phosphatidylinositol 4-kinase (PI4K), a membrane-associated PI kinase, plays a central role in cell signalling and trafficking.
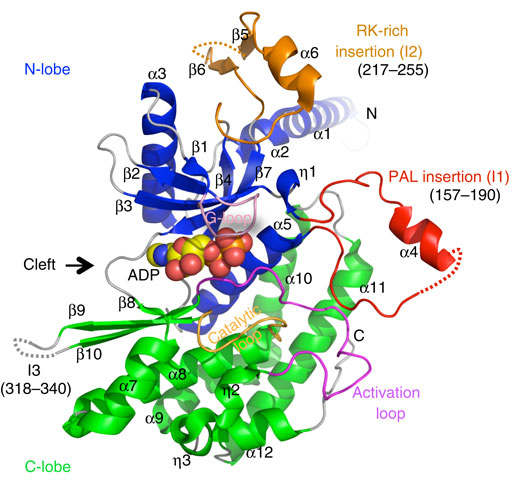
Figure 1: Overall structure of the PI4K catalytic domain. Untraced segments are depicted with dashed lines. The G-loop, catalytic loop and activation loop are coloured in pink, bright orange and magenta, respectively. The palmitoylation insertion (PAL insertion, I1) and RK-rich insertion (I2) are coloured in red and gold, respectively. ADP is shown in a sphere model and is coloured according to its atoms (carbon, yellow; oxygen, red; nitrogen, blue; phosphorus, orange).
The PI4K family includes type II PI4Ks (PI4KIIs) and type III PI4Ks (PI4KIIIs). Unlike PI4KIIIs that belong to the PI 3/4 kinase family and are structurally similar to PI3Ks, PI4KIIs have a catalytic region that differs substantially from the one found in PI3Ks. PI4K is insensitive to inhibitors of PI3Ks, but can be inhibited by ADP, AMP and adenosine. Although many specific high-quality isoform-selective inhibitors of PI3Ks have been developed based on the crystal structures of PI3Ks and some of them, such as PX866, GS-1101 and PI-103, have already been tested in clinical trials, no specific inhibitor for PI4K is currently available.
Type II PI4K, the most abundant PI4K in mammalian cells and mainly localized in the trans-Golgi network (TGN) and endosomes, produces more than half of the Golgi phosphatidylinositol 4-phosphate and is involved in many different cell pathways including PI(4,5)P2 synthesis, membrane trafficking (regulating endosomal sorting of signalling receptors and promoting adaptor protein recruitment to endosomes and TGN), signal transduction (Wnt and HER-2 signalling pathways), phagocytosis and the exo-endocytic cycle of synaptic vesicles. Its dysfunction contributes to tumour growth, spastic paraplegia, Gaucher's disease and Alzheimer's disease, suggesting PI4K as a drug target. Its kinase activity critically depends on palmitoylation of its cysteine-rich motif (-CCPCC-) and is modulated by the membrane environment. However it is still unclear how PI4K senses membrane environment and performs its functions.
Based on newly identified structure of PI4K, six 0.27-million atom, 1-us all-atom simulations were performed. Combining biochemical data and MD simulations, the researchers concluded that PI4K's palmitoylation does not change the strength of its membrane binding significantly, but rather modulates the fluctuations of the protein conformation and, thereby, regulates its kinase activity via tuning the PI-binding pocket. The results suggested the importance of membrane environment for PI4K activity, and provided molecular insight into how the kinase activity of PI4K can be regulated via changing membrane organization.
In this website, we briefly describe the role of PI4K control in lipid signaling. For more details, take a look at the publications and links on this web site or contact the investigators listed below.
The important role of the palmitoylation insertion
Full-length human PI4K contains a Pro-rich amino-terminal domain (1 to 93) and a carboxy-terminal catalytic domain (94 to 479), the latter including a palmitoylation motif -CCPCC- (Fig. 1). The resolved structure comprises an N-terminal flexible helix alpha1 (78 to 94) and a catalytic domain that is divided into an N-lobe (95 to 265) and a C-lobe (266 to 453). The N-lobe contains a five-stranded antiparallel beta-sheet core (beta1-beta2-beta3-beta7-beta4) flanked by a helical hairpin (alpha2 and alpha3) on one side and helix alpha5 on the other. The C-lobe contains the helices alpha8, eta2, eta3, alpha9, alpha10, alpha11 and alpha12, which form a helical bundle together with N-lobe helix alpha5. The helical bundle is flanked by three beta-strands (beta8, beta9 and beta10) and helix alpha7.
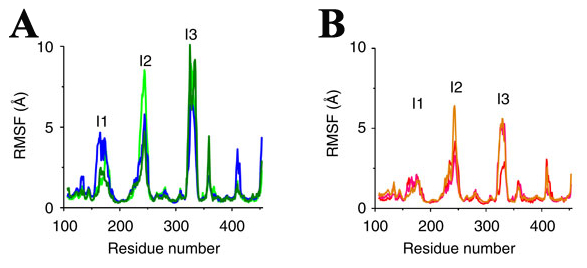
Figure 2: Conformational fluctuations of PI4K without palmitoylation in (a) and with palmitoylation in (b). Blue, green and dark green curves in (a) represent the RMSF statistics from three independent simulations of PI4K without palmitoylation. Orange, red and pink curves in (b) represent three independent simulation of palmitoylated PI4K.
PI4K has a specific cysteine-rich motif (-CCPCC-); its kinase activity critically depends on palmitoylation of the cysteine-rich motif and is modulated by cholesterol, amyloid-beta peptide, mastoparan, 25-hydroxycholesterol and ginsenoside (20S)Rg3 via changing the membrane environment as suggested previously.
The possible regulation mechanism via PI4K's membrane interaction was further investigated based on the MD simulation data (Fig. 2, 3). We monitored the relative height of PI4K's centre of mass with respect to the lipid membrane (Fig. 3b) and the membrane-insertion depth of the palmitoylation insertion (residue 166 to 179) (Fig. 3c). It is found that palmitoylation has little effect on the relative height and membrane-insertion depth. However, there is a significant structural fluctuation change as reflected by the root mean square fluctuation of the proteins. With reference to the protein conformation at t=300 ns, in the following 700 ns simulations, PI4K exhibits large conformational fluctuation around three structural insertions, the palmitoylation insertion (I1), RK-rich insertion (I2) and insertion I3. Such large conformational fluctuations were not observed in the membrane-free simulation (Fig. 3d), suggesting that they originate from PI4K's interaction with the membrane. In additional, such conformational fluctuations are reduced by 50% for the palmitoylated protein, especially at the palmitoylation insertion region (Fig. 2).
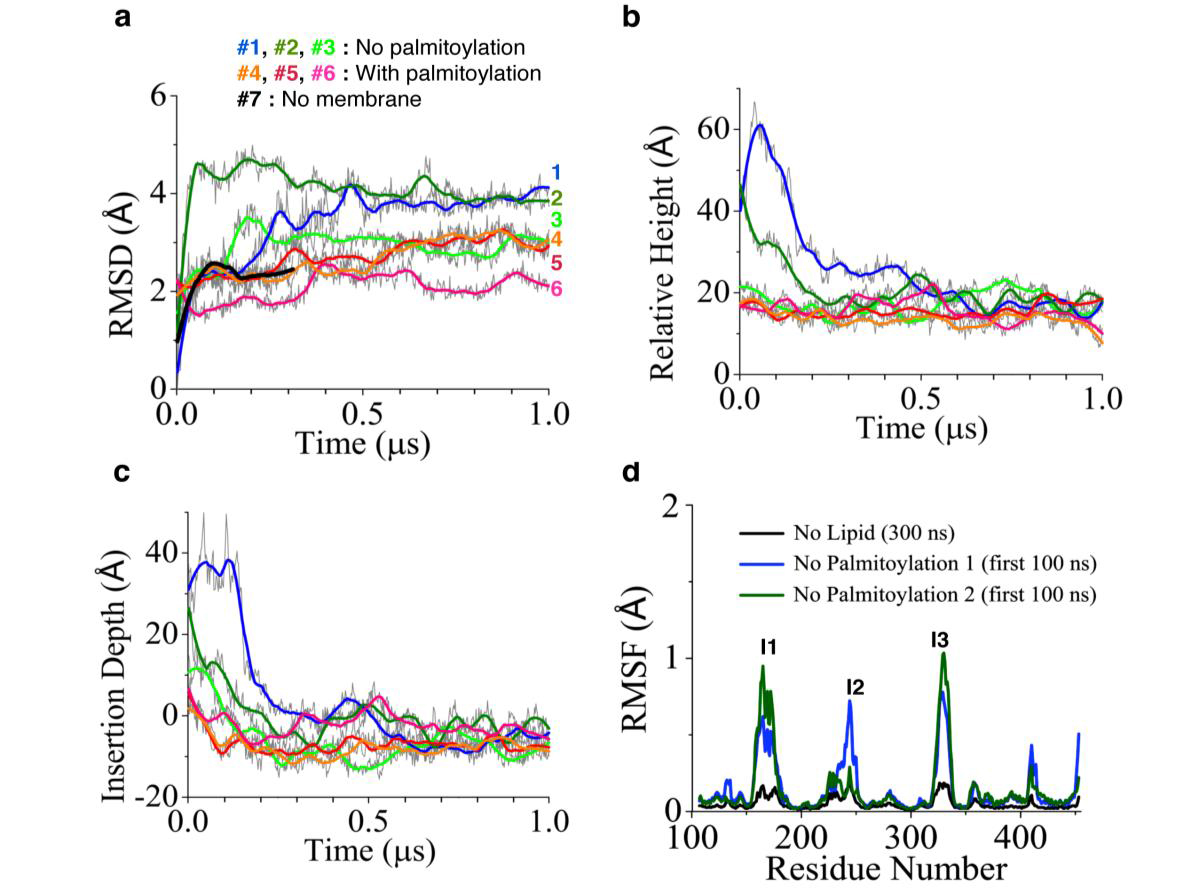
Figure 3: Structural characterization of PI4K in molecular dynamics simulations. Time evolution of (a) RMSD, (b) Relative height, (c) Insertion depth of PI4K relative to the crystal structure, with #1, #2 and #3 representing simulations of non-palmitoylated proteins, #4, #5 and #6 simulations of palmitoylated ones and #7 a simulation of membrane-free non-palmitoylated PI4K. (d) Conformational fluctuations of non-palmitoylated PI4K in the absence of interactions with a membrane. The black curve represents the membrane-freee simulations of PI4K; blue and dar-green curves were obtained from the first 100 ns of non-palmitoylated PI4K simulation in the presence of a membrane.
Based on the analysis of biochemical data and MD simulations (Fig. 2, 3), we conclude that PI4K's palmitoylation does not change the strength of its membrane binding significantly, but rather modulates the fluctuations of the protein conformation and, thereby, regulates its kinase activity via tuning the PI-binding pocket. Therefore, reduced fluctuation due to palmitoylation insertion (Fig. 2) would increase the stability of the PI-binding pocket and, thereby, increase catalytic efficiency, resulting in net increase of kinase activity. Our observations suggested the importance of membrane environment for PI4K activity, which was also proposed from previous studies based on the Golgi/endosomal pool.
Publications
Investigators
Related TCB Group Projects
- F-BAR domains
- Membrane Sculpting by BAR domains
- Intrinsic Curvature Properties of Photosynthetic Proteins in Chromatophores
Page created and maintained by Hang Yu.


