Biological Photoreceptors
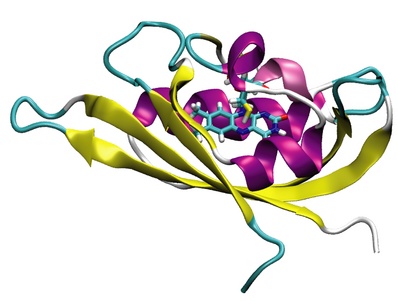 Figure 1: Molecular view of the LOV domain of C. reinhardtii. The protein is
shown in Cartoon representation; the chromophore FMN and the catalytic
cysteine residue are shown in Licorice representation.
Figure 1: Molecular view of the LOV domain of C. reinhardtii. The protein is
shown in Cartoon representation; the chromophore FMN and the catalytic
cysteine residue are shown in Licorice representation.
The conversion of sunlight into useful energy by plants and other organisms is of utmost importance for all life. This biological conversion process is called photosynthesis and transforms the electromagnetic energy of sunlight into chemical energy storage by synthesizing organic compounds (carbohydrates) from inorganic ones (carbon dioxide and water). Sunlight not only powers the conversion to chemical energy, it also provides important cues needed to regulate optimal exposure to the sunlight. For this purpose these organisms use proteins known as photoreceptors as their "eyes". Presently, three different types of photoreceptor proteins have been identified in biological systems, the red-light sensitive phytochromes, and the blue-light sensitive cryptochromes and phototropins. The latter are responsible for a wide variety of light-induced responses and have been the subject of a recent computational study employing combined quantum mechanical/molecular mechanical (QM/MM) simulations. Phototropins mediate, for example, phototropism, the light induced growth of plants toward light. Phototropism allows plants to optimize their utilization of the available sunlight, and, importantly, also enables them to avoid possible damage by overexposure to light. Other examples of light induced responses include stomatal opening and closing (the opening and closing of pores in, e.g., plant leaves to regulate the evaporation of water), chloroplast relocation (the movement of chloroplasts inside cells toward areas of higher light intensity under weak light conditions and away from high intesity areas in the opposite case), and gametogenesis (the formation of germ cells during meiosis).
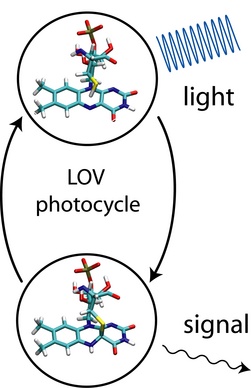
Figure 2: Schematic view of LOV photocycle.
Phototropins
Phototropin proteins consist of two main units: (1) a C-terminal serine-threonine kinase domain which is responsible for the phosphorylation of an yet unknown downstream signaling component; (2) coupled to it at the N-terminal side of the protein two so-called Light, Oxygen, and Voltage (LOV) sensitive domains, LOV1 and LOV2. Figure 1 depicts the structure of the LOV1 domain from a phototropin of the algae C. reinhardtii. The LOV domains are the actual light absorbing units in phototropins and are part of the well-characterized PER-ARNDT-SIM (PAS) family of receptor proteins. The chromophore responsible for light absorption in LOV is the molecule flavin-mononucleotide (FMN), which normally functions as a co-enzyme in redox processes during energy metabolism and which is derived from riboflavin, better known as vitamin B2. Figure 1 shows that FMN is non-covalently bound in the core of the LOV domain; the top part of Figure 2 provides a close up view of FMN itself. The photodynamics of LOV domains have been studied experimentally using IR and EPR spectroscopy. Furthermore, the crystal structures of LOV domains of several species have recently been determined at high resolution. These studies revealed that the initial absorption of blue light by the non-covalently bound chromophore FMN promotes the system into an excited triplet state. This triplet state then initiates the formation of a covalent flavin-cysteinyl adduct state between FMN and a cysteine residue of the LOV protein, which is located close to the flavin ring (see Figure 2, bottom). The LOV domain in its flavin-cysteinyl adduct state then transmits a signal to the attached kinase domain, thereby, initiating signaling of the whole phototropin protein. The conversion back to the initial non-signaling, non-adduct state (see Figure 2, top) is thermally activated and happens on a timescale of ~100s. Figure 2 gives a schematic overview of the flavin-cysteinyl bond formation and bond breaking process that occurs during the photocycle.
Despite the availability of spectroscopic and structural data on LOV’s photocycle science still lacks a detailed understanding of the underlying events, particularly on a microscopic and electronic level. For example, it is not known how the bond formation step in the excited triplet state is initiated. Several possible mechanism have been proposed in the literature. Furthermore, we currently do not know how signaling takes place after the flavin-cysteinyl bond between FMN and LOV has been formed. From X-ray crystallographic structures, both the inactive and the flavin-cysteinyl adduct state have been found to be very similar, suggesting a change in protein dynamics rather than conformation as the relevant switch mechanism.
When Light Falls in LOV
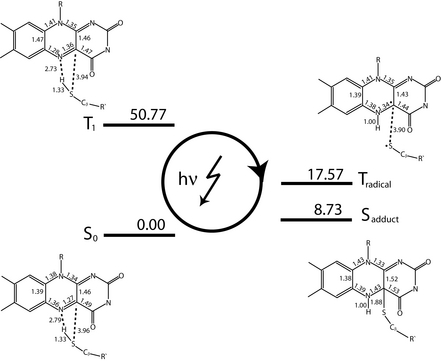
Figure 3: LOV photocycle. Shown is a schematic representation of the photocyle determined via QM/MM simulations.
To investigate these questions in more detail we have employed QM/MM simulations to study on an electronic level the events during photoexcitation of the LOV1 domain from the algae C. reinhardtii. Here, a quantum mechanical simulation method was used to explicitly treat the electronic degrees of freedom of those parts of the system that are directly involved in photoactivation. The remainder of the protein as well as the solvent environment was treated using a classical molecular mechanics forcefield. Our simulations allowed the characterization of the complete LOV photocycle on an electronic level as well as the identification of several intermediate states in the excited triplet state. Figure 3 shows several states of LOV domain’s photocycle identified by our simulations together with their energies and geometries. The initial triplet state T1, created via absorption of a blue-light photon, converts into a second triplet state, Tradical (see Figure 3). Tradical was found to be a neutral radical-pair state evolving from T1 via transfer of an hydrogen atom. Furthermore, our simulations showed that Tradical converts to the covalent flavin-cysteinyl state Sadduct (c.f. Figure 3) via a crossing between the triplet and singlet potential energy surfaces. Hence, our simulations provide clear evidence for a covalent flavin-cysteinyl bond formation between the chromophore FMN and LOV via a neutral radical-pair mechanism.
LOV Domain Activation
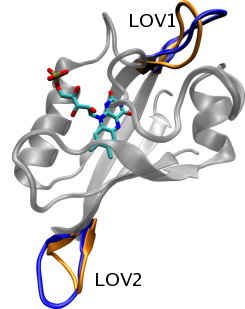
Figure 4: Regions of high mobility in LOV1 and LOV2, as discussed in the text. In both cases, the blue conformation is the inactive state, and orange the photoactivated state.
After understanding the electronic-level mechanism of the LOV domain photoreaction, it becomes important to determine how this chemical change leads to phototropin kinase activation. X-ray crystal structures comparing the light and dark states of LOV1 and LOV2 showed only very minor changes in average structure after the photoreaction, indicating that the changes triggered are likely dynamic in nature. To determine how the LOV domain photoreaction might trigger activation, we performed long-timescale molecular dynamics simulations on both the dark (inactive) and light (active) states of LOV1 and LOV2. Analysis of these simulations were in agreement with crystal structures in indicating that the average structure of the protein does not change after photoactivation; however, regions of unusual mobility were observed in both LOV1 and LOV2 (see Figure 4). In the case of LOV1, a salt bridge which was broken prior to photoactivation was found to form in the light state, changing the conformation of a loop and removing a region of negative electrostatic potential from the protein's surface. In LOV2, the same salt bridge was formed in both the dark and light states, but a loop, on the opposite end of the protein undergoes a conformational shift; this loop is, furthermore, close to a highly conserved helix thought to be involved in LOV2 activation. Thus, our MD simulations revealed a significant difference in the dynamic changes in LOV1 and LOV2 after photoactivation, and provided potential mechanisms for modulation of kinase activity in both cases.
Publications
Resources
We have developed several generations of parameters for flavin mononucleotide (FMN) and the FMN-cysteine photoadduct occuring in the LOV domain. The most recent version, used for the 2011 and 2013 papers listed above, is described in our recent publication and can be downloaded here.Important technical note regarding FMN parameters: The first generation of our FMN/FMC parameters, included with our 2006 article, was developed for NAMD 2.5 and earlier; these versions of NAMD use a sign convention on the offset of dihedral angles that differs from CHARMM and more recent versions of NAMD. Users wishing to apply these paramaters with later versions of NAMD should contact Peter Freddolino for more information. Unless you have a specific reason for wanting the older parameters, we highly recommend the more recent parameter set linked above, which has substantially improved partial charges (and uses the dihedral angle convention of NAMD 2.6b1+ and CHARMM).
Investigators
Other QM/MM projects
Acknowledgment
This material is based upon work supported by the National Science Foundation under Grant No. 0234938. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.


