Comparing Models of the Visual Cortex
Comparing Models of the Visual Cortex
Introduction
One goal of our research is to use cameras for robot control in a biologically inspired manner by integrating units similar to cortical visual and motor maps. Important for this goal is an understanding of the principles underlying the structure of visual maps. Many different models have been proposed during the past two decades. We undertook a critical evaluation of the most prominent and successful of the alternative approaches. Our goals were to exclude unsatisfactory approaches, reveal common principles underlying seemingly different models, and determine which quantities are sensible to measure in order to decide between hypotheses. This work has been carried out by Ed Erwin, a graduate student in Chemical Physics who joined the group four years ago, in collaboration with Klaus Obermayer, a past graduate student in our group and currently at the University of Bielefeld, Germany. Experimental data for the comparisons were available through a collaboration with Gary Blasdel of Harvard Medical School.
Description
Many cells in the mammalian primary visual cortex are selective to certain features in visual input. For example, they respond better to stimulation of one eye over the other, and usually respond more strongly to bars or gratings of a particular orientation. The figure shows the lateral spatial pattern of ocular dominance and orientation selectivity in the striate cortex of an adult macaque. Examples are shown of several elements of the lateral organization which have been termed (1) singularities, (2) linear zones, (3) saddle points, and (4) fractures. These and similar large, high-resolution pictures have led to a greatly improved characterization of striate cortical organization in the macaque.
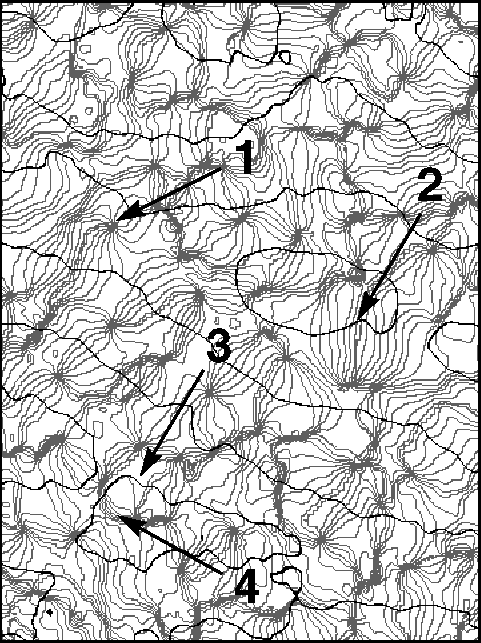
The lateral spatial pattern of orientation preference and ocular dominance in the striate cortex of an adult macaque as revealed by optical imaging. The figure shows a 4.1mm x 3.0 mm surface region located near the border between cortical areas 17 and 18 and close to the midline. Black contours separate bands of opposite eye dominance. Light gray iso-orientation contour lines separate regions responsive to orientations differing by 11.25 degrees. The medium gray contour represents the preferred orientation 0 degree. Arrows indicate (1) singularities, (2) linear zones, (3) saddle points, and (4) fractures.
Many models for the structure and formation of orientation and ocular dominance maps have been proposed. Although seemingly based on different assumptions, most produce maps which visually resemble the experimentally obtained maps. To sort through the conflicting claims, we extended and analysed the most prominent of the previously proposed models and compared their predictions with the experimental data.
For convenience we grouped the major models into classes and types based on similarities in goals or implementation. The two main classes are pattern models and developmental models. Pattern models attempt to provide a characterization of map patterns in schematic drawings or simple equations. They include both structural and spectral models. Developmental models give more attention to proposed mechanisms of pattern formation. Their pattern-generating equations are meant to correspond more closely to actual physiological processes. Developmental models include correlation-based learning, competitive Hebbian models, and several others.
This study revealed that several pattern models, as well as an entire class of developmental correlation-based learning models, are incompatible with experimental data, whereas competitive Hebbian models and several particular pattern models are in accordance with the experimental findings. Furthermore, we found that despite apparent differences, most models are based on similar principles and consequently make similar predictions.
The main results are as follows: Several pattern models and several developmental models generate maps which closely match the experimentally observed map patterns. Among the pattern models, the spectral models perform better than the previously proposed structural models, mainly because they account for global disorder and for the coexistence of linear zones and singularities. The filtered noise approach for orientation selectivity and for ocular dominance captures most of the important features of the individual maps, except for the high degree of feature selectivity which is observed in the macaque. Models by Swindale provide the currently best description of the patterns found in the macaque and can additionally account for the correlations between the orientation and ocular dominance maps. The close agreement, however, is reached at the expense of extreme simplification of the biological processes.
Among the developmental models, our analysis showed that the correlation-based learning models, which involve Hebbian learning but which include only linear intra-cortical interactions, perform well for ocular dominance. When applied to the formation of orientation maps, these models fail to predict the existence of linear zones and wrongly predict correlations between cells' orientation preferences and their spatial relationship in cortex. Correlation-based learning models have led to valuable insight into the role of Hebbian learning in receptive field development. However, to be applicable to cortical map formation, the nature of the lateral interaction must be changed.
Competitive Hebbian models --- which are based on non-linear lateral interactions --- lead to the currently best description of the observed patterns from a developmental model. These models attempt to describe the developmental process on a mesoscopic level, spatially as well as temporally, which has the advantage that the level of description matches the resolution of the experimental data. These models do not involve the microscopic concepts neuron, synapse and spike, which makes it somewhat more difficult to relate model predictions to experimental data.
Improvements on present models will require additional data on the spatial patterns. More stringent tests of the postulated mechanisms of activity-dependent neural development must rely on experiments which monitor the actual time-course of pattern formation, and which study pattern development under experimentally modified conditions (deprivation experiments). Mechanisms of visual cortex development should be fairly universal. Thus, any model of value should, additionally, be able to account for inter-species variations. Finally, one would like to have relatively simple models which make predictions about several aspects of cortical organization, such as receptive field location, color selectivity, receptive field sub-fields, and spatial phase. Correlations between maps of different features predicted by such models could be tested in suitably designed experiments.


