Respiration and photosynthesis are ubiquitous bioenergetic processes for harvesting and converting energy in biological cells. Many key proteins and physical processes are common to both processes, featuring organelle-scale arrays of membrane protein
complexes. Dysfunction of the repiratory complex is associated with several pathologies, including Parkinson's disease,
malaria, and breast cancer. Here, molecular modeling techniques from single-protein to organelle scales
are employed to study
(1) energy conversion in a respiratory supercomplex, called respirasome, in mitochondria(S1) and (2) energy harvesting by a granal thylakoid membrane in chloroplasts(S2).
Innovation
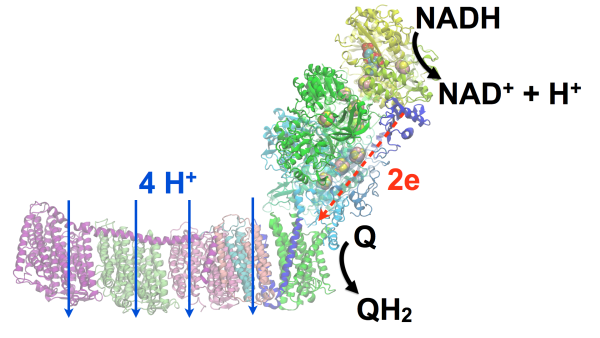
A central challenge for both (S1) and (S2) is the lack of atomic-detail structural models of the
supramolecular organization. The structural model for complex I (NADH dehydrogenase) (S1,S2) as well as the entire respirasome (S1) will be resolved
in atomic detail by extending current MDFF methods,
driving the development of autoMDFF (TRD3) protocols for the resolution of sub-5 angstrom EM maps. The granal thylakoid (S2), featuring nearly 2 billion atoms and comprising over 10,000 proteins in a 500 nm membrane domain, will be investigated through tools for cell-scale placement of macromolecules (TRD2), large system visualization (TRD2),
and the CMMxmasbuilder (TRD3) membrane building protocol
for the construction of atomic-detail organelle domains
by extending earlier work
on bacterial bioenergetic organelles. A further challenge
due to the very large system size of S2
is that most experimental laboratories
currently lack in-house visualization capabilities for viewing and analyzing systems of this size at atomic detail,
whereas the construction of
such structural models requires visual data exchange
between experimental and computational scientists.
This exchange will be facilitated using remote visualization (TRD2) tools employing GPU-clusters in supercomputer centers. A challenge for modeling S1 and S2 is the extensive range of time-scales and length-scales involved in
the energy conversion steps, which span from microseconds for proton transport processes to milliseconds for diffusion of redox
factors spread over hundreds to thousands of proteins, requiring sampling over many trajectories.
The computational demand of such modeling efforts will employ petascale/exascale computing (TRD1) capabilities. Electrostatic potentials based on all-atom structural models will be determined employing the proPka protocols with
the advanced electrostatics (TRD2) interface. A related challenge involves pH-dependent kinetics of proton transport in
complex I (NADH dehdrogenase) (S1,S2).
This transport process will be simulated using constant pH (TRD1) simulations
and QM/MM (TRD1) methods.
The challenge for modeling the slowest, rate-determining steps involve millisecond scale diffusion processes. Such processes, including electron transfer from complex III/cytochrome b6f onto complex IV (S1) or photosystem I (S2)
via carrier proteins cytochrome c/plastocyanin, will be modelled using
GPU-BD (TRD3) protocols for
atomic resolution Brownian dynamics.
Approach
The investigations described above will be performed based on expertise of the Center over the past four decades
on modeling energy conversion processes in bioenergetic
systems. Recently, the atomic detailed structure of a bacterial bioenergetic organelle, the photosynthetic chromatophore,
was determined based on AFM, cryo-EM, crystallography, and proteomics data
(Hunter)
and the overall energy conversion into ATP was modeled.
Furthermore, a model for the light-harvesting domain of the cyanobacterial thylakoid membrane, containing 96 photosystem I complexes,
was also recently constructed based on AFM data (Hunter). Following similar protocols, a granal thylakoid membrane for the plant chloroplast will be constructed for modeling the fundamental energy harvesting processes ({\bf Hunter, Blankenship) (TRD1, TRD2, TRD3). A complete respirasome model will be constructed based on crystallographic and EM data (Sazanov) employing
MDFF (TRD3). Key residues predicted during MD simulations of proton transport in complex I
and Brownian dynamics simulation of electron transport between complexes III and IV
will be confirmed by biochemical data (Gennis) (TRD2, TRD3).
Publications
Publications Database
Atomic detail visualization of photosynthetic membranes with GPU-accelerated ray tracing.
John E. Stone, Melih Sener, Kirby L. Vandivort, Angela Barragan, Abhishek Singharoy, Ivan Teo, Joao V. Ribeiro, Barry Isralewitz, Bo Liu, Boon Chong Goh, James C. Phillips, Craig MacGregor-Chatwin, Matthew P. Johnson, Lena F. Kourkoutis, C. Neil Hunter, and Klaus Schulten. Parallel Computing, 55:17-27, 2016.
Quantum biology of retinal.
Shigehiko Hayashi and Klaus Schulten. In Masoud Mohseni, Yasser Omar, Greg Engel, and Martin B. Plenio, editors, Quantum Effects in Biology, pp. 237-263. Cambridge University Press, 2014.
Visualization of energy conversion processes in a light harvesting organelle at atomic detail.
Melih Sener, John E. Stone, Angela Barragan, Abhishek Singharoy, Ivan Teo, Kirby L. Vandivort, Barry Isralewitz, Bo Liu, Boon Chong Goh, James C. Phillips, Lena F. Kourkoutis, C. Neil Hunter, and Klaus Schulten. In Proceedings of the International Conference on High Performance Computing, Networking, Storage and Analysis, SC '14. IEEE Press, 2014. (4 pages).
Ultrastable cellulosome-adhesion complex tightens under load.
Constantin Schoeler, Klara H. Malinowska, Rafael C. Bernardi, Lukas F. Milles, Markus A. Jobst, Ellis Durner, Wolfgang Ott, Daniel B. Fried, Edward A. Bayer, Klaus Schulten, Hermann E. Gaub, and Michael A. Nash. Nature Communications, 5:5635, 2014.
The FMO complex in a glycerol-water mixture.
Mortaza Aghtar, Johan Strümpfer, Carsten Olbrich, Klaus Schulten, and Ulrich Kleinekathoefer. Journal of Physical Chemistry B, 117:7157-7163, 2013.
Molecular dynamics methods for bioelectronic systems in photosynthesis.
Ioan Kosztin and Klaus Schulten. In Thijs Aartsma and Joerg Matysik, editors, Biophysical Techniques in Photosynthesis II, volume 26 of Advances in Photosynthesis and Respiration, pp. 445-464. Springer, Dordrecht, 2008.